L’exposition prolongée aux rayonnements ultraviolets représente l’une des principales menaces pour la santé cutanée. Bien que les effets immédiats comme les coups de soleil soient visibles, les conséquences à long terme s’avèrent bien plus préoccupantes et irréversibles. Les radiations UV provoquent des altérations moléculaires profondes qui s’accumulent silencieusement au fil des années, compromettant l’intégrité structurelle et fonctionnelle de la peau. Ces dommages progressifs engendrent des pathologies graves allant du vieillissement prématuré aux cancers cutanés les plus agressifs.
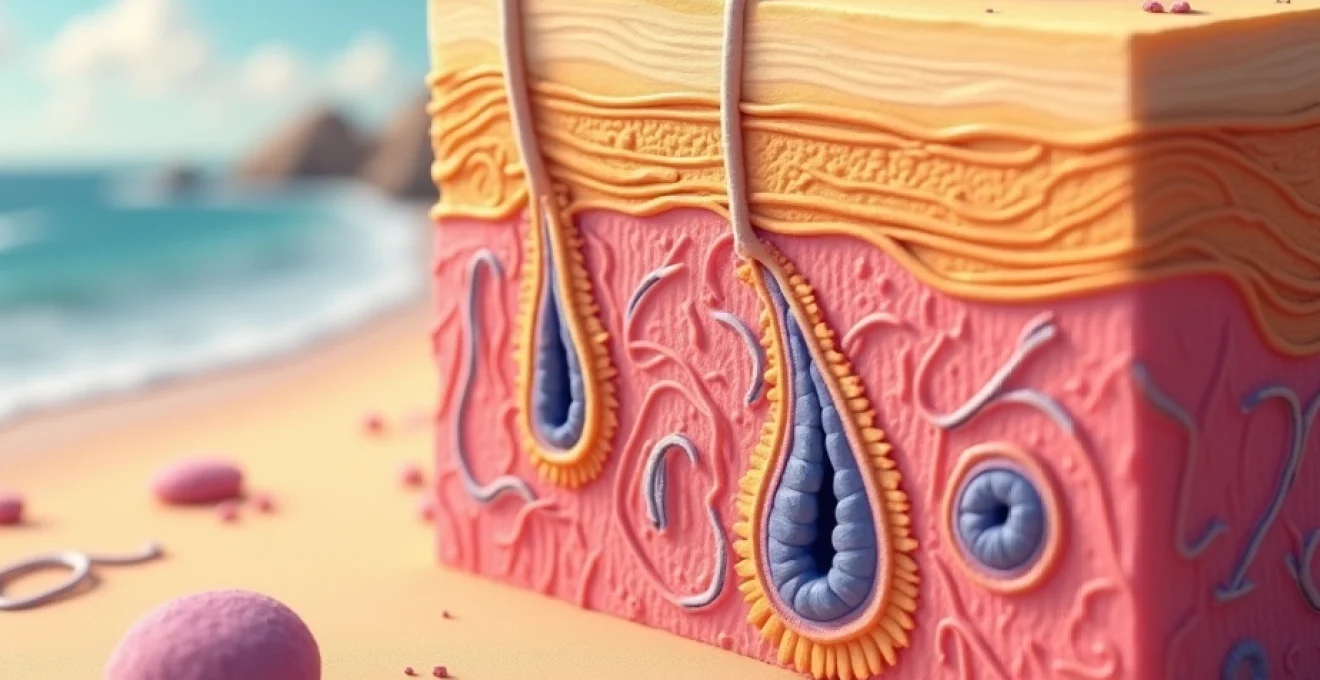
Mécanismes biophysiques de la pénétration des radiations ultraviolettes dans l’épiderme et le derme
La compréhension des mécanismes de pénétration des rayons UV constitue la base fondamentale pour saisir leurs effets délétères à long terme. Lorsque les radiations ultraviolettes atteignent la surface cutanée, elles interagissent différemment selon leur longueur d’onde et leur énergie photonique. Cette interaction initiale détermine la profondeur de pénétration et l’ampleur des dommages cellulaires qui s’ensuivent.
Absorption différentielle des UV-A (320-400 nm) et UV-B (280-320 nm) par les chromophores cutanés
Les UV-A et UV-B présentent des profils de pénétration radicalement différents dans les tissus cutanés. Les UV-A, moins énergétiques mais plus pénétrants, traversent facilement l’épiderme pour atteindre le derme profond et même l’hypoderme. Cette capacité de pénétration explique leur rôle prépondérant dans le photovieillissement et l’altération de la matrice extracellulaire. Représentant 95% des UV atteignant la surface terrestre , ils maintiennent une intensité relativement constante tout au long de la journée et traversent partiellement la couverture nuageuse.
Les UV-B, plus énergétiques mais moins pénétrants, sont majoritairement absorbés par l’épiderme, particulièrement par les kératinocytes des couches superficielles. Leur absorption par l’ADN cellulaire génère des lésions directes responsables de la mutagenèse. Les chromophores cutanés, incluant les acides nucléiques, les protéines aromatiques et la mélanine, constituent les cibles primaires de cette absorption photonique.
Interaction photochimique entre les photons UV et l’ADN mitochondrial des kératinocytes
L’interaction entre les photons UV et l’ADN cellulaire représente l’événement moléculaire le plus critique dans l’initiation des dommages à long terme. Les UV-B, absorbés directement par les bases pyrimidiques de l’ADN, induisent la formation de dimères cyclobutylpyrimidiques et de photoproduits 6-4. Ces lésions perturbent la structure hélicoïdale de l’ADN et compromettent la fidélité de la réplication cellulaire.
L’ADN mitochondrial des kératinocytes s’avère particulièrement vulnérable aux dommages UV en raison de sa proximité avec les systèmes de production d’espèces réactives oxygénées. Cette vulnérabilité mitochondriale amplifie les effets du stress oxydatif et compromet le métabolisme énergétique cellulaire. La réparation de ces lésions mitochondriales s’effectue moins efficacement que pour l’ADN nucléaire, entraînant une accumulation progressive de mutations délétères.
Altération de la barrière lipidique intercornéocytaire par photooxydation des céramides
La fonction barrière de la peau dépend essentiellement de l’intégrité des lipides intercornéocytaires, notamment les céramides, le cholestérol et les acides gras libres. L’exposition UV chronique provoque la photooxydation de ces composants lipidiques, altérant leur organisation lamellaire et compromettant leur fonction d’étanchéité. Cette dégradation lipidique entraîne une augmentation de la perte transépidermique en eau et une vulnérabilité accrue aux agressions extérieures.
Les céramides, constituants majoritaires du ciment lipidique intercornéocytaire, subissent une peroxydation lipidique sous l’effet des espèces réactives générées par les UV. Cette altération structurelle compromet la cohésion de la couche cornée et favorise la déshydratation cutanée. La photooxydation des céramides contribue directement à l’apparition des signes cliniques de photovieillissement , incluant la rugosité, la sécheresse et l’épaississement irrégulier de l’épiderme.
Formation de dimères de thymine et de produits de photooxydation dans les cellules basales
La formation de dimères de thymine constitue l’une des lésions ADN les plus caractéristiques de l’exposition UV-B. Ces lésions se forment lorsque deux thymines adjacentes s’associent covalemment, créant une distorsion structurelle majeure de la double hélice. Dans les cellules basales, particulièrement actives en division cellulaire, ces dimères compromettent la progression de la fourche de réplication et induisent des mutations de substitution.
Les produits de photooxydation, générés par l’action indirecte des UV via la production d’espèces réactives oxygénées, créent un spectre plus large de lésions ADN. La 8-oxoguanine, principal adduit oxydatif de l’ADN, s’accumule progressivement dans les cellules basales exposées chroniquement aux UV. Ces lésions oxydatives échappent partiellement aux systèmes de réparation cellulaire et constituent des sites préférentiels de mutagenèse spontanée.
Photocarcinogenèse cutanée : progression moléculaire vers les néoplasies malignes
La photocarcinogenèse représente un processus complexe et multi-étapes où l’accumulation de mutations induites par les UV transforme progressivement les cellules normales en cellules néoplasiques. Ce processus implique l’altération séquentielle de gènes suppresseurs de tumeurs et d’oncogènes, créant un microenvironnement propice au développement tumoral. La compréhension de ces mécanismes moléculaires éclaire la progression inexorable vers les formes les plus agressives de cancers cutanés.
Mutations oncogéniques induites par les UV dans les gènes p53 et PTCH1
Le gène p53, surnommé « gardien du génome », constitue la cible privilégiée des mutations induites par les UV. Plus de 90% des carcinomes spinocellulaires présentent des mutations de p53, principalement des transitions C→T caractéristiques de l’exposition UV-B. Ces mutations inactivent la fonction suppresseur de tumeur de p53, permettant la survie et la prolifération de cellules porteuses de lésions ADN critiques.
Le gène PTCH1, régulateur essentiel de la voie de signalisation Hedgehog, subit également des mutations spécifiques sous l’exposition UV. L’inactivation de PTCH1 libère la signalisation oncogénique de cette voie , favorisant la prolifération cellulaire incontrôlée caractéristique des carcinomes basocellulaires. La signature mutationnelle UV dans PTCH1 confirme l’étiologie solaire de ces tumeurs et explique leur prédilection pour les zones photo-exposées.
Développement des carcinomes basocellulaires nodulaires et sclérodermiformes
Les carcinomes basocellulaires représentent les tumeurs cutanées les plus fréquentes, avec une incidence directement corrélée à l’exposition UV cumulative. La forme nodulaire, caractérisée par une prolifération de cellules basaloïdes organisées en nodules bien délimités, évolue lentement et reste généralement localisée. Cette variante histologique présente un stroma fibromyxoïde riche en mucine et une vascularisation périphérique caractéristique.
La forme sclérodermiforme, plus agressive, se caractérise par une croissance infiltrante avec des prolongements tumoraux étendus dans le derme. Cette variante présente un potentiel invasif supérieur et une tendance récidivante marquée en cas d’exérèse incomplète. Le carcinome basocellulaire sclérodermiforme nécessite des marges d’exérèse plus larges en raison de ses extensions microscopiques imprévisibles dans les tissus adjacents.
Évolution des carcinomes épidermoïdes in situ vers les formes invasives
Les carcinomes épidermoïdes suivent une progression histologique bien documentée, débutant par des kératoses actiniques puis évoluant vers des formes in situ avant d’acquérir un potentiel invasif. Cette progression reflète l’accumulation séquentielle de mutations dans les gènes régulateurs du cycle cellulaire et de l’adhésion intercellulaire. Les formes in situ, confinées à l’épiderme, présentent une dysplasie kératinocytaire complète mais conservent l’intégrité de la membrane basale.
La transition vers l’invasion dermique implique l’activation de métalloprotéases matricielles et la dégradation de la membrane basale. Cette étape critique transforme une lésion localisée en tumeur potentiellement métastatique. Le potentiel métastatique des carcinomes épidermoïdes photo-induits augmente significativement avec l’épaisseur tumorale et le degré de différenciation histologique, justifiant une surveillance oncologique rigoureuse.
Transformation maligne des mélanocytes : mélanome superficiel extensif et mélanome nodulaire
La transformation maligne des mélanocytes représente la conséquence la plus redoutable de l’exposition UV intermittente et intense. Le mélanome superficiel extensif, forme la plus fréquente, se caractérise par une phase de croissance horizontale prolongée avant l’acquisition d’un potentiel invasif vertical. Cette progression biphasique offre une fenêtre thérapeutique cruciale pour un diagnostic précoce et un traitement curatif.
Le mélanome nodulaire, forme plus agressive, présente d’emblée une croissance verticale invasive sans phase horizontale préalable. Cette variante histologique associe un pronostic plus sombre en raison de sa capacité métastatique précoce et de son diagnostic souvent tardif. L’exposition UV intermittente et les coups de soleil sévères durant l’enfance constituent les facteurs de risque majeurs pour le développement de ces formes agressives de mélanome.
Photovieillissement prématuré : dégradation structurelle de la matrice extracellulaire
Le photovieillissement constitue un processus distinct du vieillissement chronologique, caractérisé par des altérations structurelles spécifiques de la peau photo-exposée. Ce phénomène résulte de l’action cumulative des radiations UV sur les composants de la matrice extracellulaire dermique, entraînant une désorganisation progressive de l’architecture tissulaire. Les manifestations cliniques du photovieillissement incluent les rides profondes, la perte d’élasticité, l’hyperpigmentation irrégulière et la télangiectasie.
Activation des métalloprotéases matricielles MMP-1, MMP-3 et MMP-9 par stress oxydatif
L’exposition UV déclenche une cascade de signalisation cellulaire aboutissant à la surexpression de métalloprotéases matricielles (MMP). Les MMP-1, MMP-3 et MMP-9 représentent les enzymes clés de cette dégradation matricielle, ciblant respectivement le collagène fibrillaire, les protéoglycanes et la gélatine. L’activation de ces enzymes résulte principalement du stress oxydatif induit par les espèces réactives oxygénées générées lors de l’exposition UV.
La régulation de ces MMP implique la voie de signalisation AP-1 (Activator Protein-1), activée par les kinases JNK et p38 en réponse au stress UV. Cette suractivation enzymatique crée un déséquilibre entre la synthèse et la dégradation matricielle , favorisant l’accumulation de produits de dégradation anormaux et la désorganisation de l’architecture dermique. L’inhibition naturelle de ces MMP par les TIMP (Tissue Inhibitors of Metalloproteinases) s’avère insuffisante pour contrebalancer leur surexpression chronique.
Fragmentation enzymatique du collagène de type I et de l’élastine dermique
Le collagène de type I, constituant majoritaire de la matrice dermique, subit une fragmentation progressive sous l’action des MMP-1 et MMP-8 activées par l’exposition UV chronique. Cette dégradation collagénique compromet la résistance mécanique et l’élasticité cutanée, se traduisant cliniquement par l’apparition de rides et l’affaissement tissulaire. Les fragments collagéniques générés exercent un rétrocontrôle négatif sur la synthèse de nouveau collagène par les fibroblastes.
L’élastine dermique, responsable de l’élasticité cutanée, subit une dégradation similaire par les élastases activées par le stress UV. Cette protéine, caractérisée par sa grande stabilité métabolique, ne peut être efficacement renouvelée chez l’adulte. La perte d’élastine constitue donc un processus irréversible qui s’accumule avec l’âge et l’exposition solaire, expliquant la perte progressive d’élasticité cutanée observée dans le photovieillissement.
Formation d’élastose solaire et d’accumulation de matériel élastotique anormal
L’élastose solaire représente l’une des caractéristiques histologiques les plus spécifiques du photovieillissement. Ce phénomène se caractérise par l’accumulation de matériel élastotique anormal dans le derme papillaire et réticulaire superficiel. Ce matériel, composé d’élastine dégradée et de fibres élastiques dystrophiques, confère à la peau photo-vieillie son aspect clinique caractéristique de « cuir tanné ».
La formation d’élastose solaire résulte d’un processus complexe impliquant la dégradation de l’élastine normale et la synthèse aber
rante d’élastine modifiée par les fibroblastes activés par l’exposition UV chronique. Cette néosynthèse d’élastine anormale ne compense pas la perte fonctionnelle de l’élastine native et contribue paradoxalement à la rigidification tissulaire. L’élastose solaire constitue un marqueur histologique pathognomonique de l’exposition UV chronique et corrèle directement avec l’intensité et la durée de l’exposition solaire cumulative.
Dysfonctionnement des fibroblastes et diminution de la synthèse de glycosaminoglycanes
Les fibroblastes dermiques, cellules responsables de la synthèse et du maintien de la matrice extracellulaire, subissent des altérations fonctionnelles majeures sous l’effet de l’exposition UV chronique. Ces cellules présentent une diminution significative de leur activité biosynthétique, particulièrement concernant la production de collagène de type I et III. Cette dysfonction fibroblastique résulte de l’accumulation de dommages oxidatifs intracellulaires et de l’altération de voies de signalisation essentielles comme TGF-β.
La synthèse de glycosaminoglycanes, notamment l’acide hyaluronique et les chondroïtines sulfates, diminue progressivement avec l’exposition UV cumulative. Ces macromolécules jouent un rôle crucial dans l’hydratation tissulaire et la viscoélasticité dermique. La perte de glycosaminoglycanes contribue à la déshydratation cutanée et à la perte de volume caractéristique du photovieillissement. Cette diminution s’accompagne d’une modification qualitative des glycosaminoglycanes synthétisés, avec une réduction de leur poids moléculaire et de leur capacité de rétention hydrique.
Immunosuppression photo-induite et défaillance des mécanismes de surveillance cellulaire
L’exposition aux radiations ultraviolettes induit une immunosuppression locale et systémique qui compromet les mécanismes naturels de surveillance immunitaire cutanée. Cette immunosuppression photo-induite résulte de multiples mécanismes cellulaires et moléculaires qui altèrent la fonction des cellules présentatrices d’antigènes, modifient les profils cytokiniques et perturbent l’équilibre entre immunité cellulaire et humorale. Ces altérations immunitaires favorisent non seulement la carcinogenèse mais également la récurrence d’infections virales cutanées.
Les cellules de Langerhans, sentinelles immunologiques de l’épiderme, subissent une déplétion significative sous l’effet des UV-B. Cette diminution quantitative s’accompagne d’une altération fonctionnelle de leur capacité de présentation antigénique et de leur migration vers les ganglions lymphatiques régionaux. L’exposition UV induit également la production de médiateurs immunosuppresseurs comme l’IL-10 et le TNF-α, créant un microenvironnement favorable à l’échappement immunitaire des cellules précancéreuses.
La fonction des lymphocytes T effecteurs se trouve compromise par l’exposition UV chronique, avec une orientation préférentielle vers un profil Th2 immunosuppresseur au détriment de la réponse Th1 cytotoxique. Cette dysrégulation immunitaire explique l’augmentation de la susceptibilité aux infections cutanées et la diminution de l’efficacité de la surveillance anti-tumorale. L’immunosuppression photo-induite persiste plusieurs semaines après l’exposition UV, créant des fenêtres de vulnérabilité prolongées pour le développement de pathologies cutanées.
Pathologies inflammatoires chroniques déclenchées par l’exposition UV cumulative
L’exposition UV cumulative déclenche et entretient des processus inflammatoires chroniques qui contribuent significativement à la pathogenèse cutanée à long terme. Ces réactions inflammatoires, caractérisées par l’infiltration de cellules inflammatoires et la production soutenue de médiateurs pro-inflammatoires, créent un environnement tissulaire propice au développement de pathologies chroniques. L’inflammation chronique photo-induite accélère les processus de vieillissement et favorise la progression néoplasique par des mécanismes épigénétiques complexes.
La dermatite actinique chronique représente l’une des manifestations les plus caractéristiques de cette inflammation UV-induite. Cette pathologie se manifeste par un érythème persistant, une hyperkératose et une infiltration lymphocytaire dense du derme superficiel. Les patients développent une photosensibilité extrême avec extension du spectre d’action vers les longueurs d’onde visibles, créant un cercle vicieux d’exposition réduite et de photosensibilisation accrue.
L’exposition UV chronique favorise également le développement de photodermatoses immunologiques comme la lucite polymorphe et l’urticaire solaire. Ces pathologies résultent d’une sensibilisation immune aux photo-antigènes générés par l’interaction UV-protéines cutanées. La persistance de ces réactions inflammatoires contribue à l’altération de la barrière cutanée et à l’accélération des processus dégénératifs dermiques. Ces pathologies inflammatoires chroniques nécessitent une photoprotection stricte et peuvent bénéficier de traitements immunosuppresseurs topiques ou systémiques pour contrôler l’inflammation et prévenir les complications à long terme.
L’inflammation chronique photo-induite active également les voies de signalisation NF-κB et AP-1, perpétuant la production de cytokines pro-inflammatoires et de métalloprotéases matricielles. Cette activation transcriptionnelle soutenue entretient un état inflammatoire de bas grade qui accélère tous les processus dégénératifs cutanés et favorise la progression tumorale par des mécanismes de promotion et de progression néoplasique.